-
Оценка достоверности реконструированной топологии с помощью бутстреп-анализа
Работа состоит из следующих этапов:
- Создание 100 бутстреп-реплик выравнивания программой fseqboot:
|
fseqboot offsprings.fasta -auto |
- Работа с программой fdnaml
fdnaml offsprings.fseqboot -ttratio 1 -auto
|
Результатом работы программы является файл offsprings.treefile, содержащий 100 скобочных формул, соответствующих реконструкциям, сделанным по каждому из выравниваний.
- Работа с программой fconsense
fconsense
|
Полученное консенсусное дерево:
+-------------D
+-62.0-| | | +------E +-97.0-| +-47.0-| | | +------F +------| | | | +--------------------A | | | +---------------------------C | +----------------------------------B |
Сравним полученное консенсусное дерево с исходным.
| Рисунок дерева в бескорневом виде | Описание ветвей дерева как разбиения множества листьев | |
| Исходное дерево | 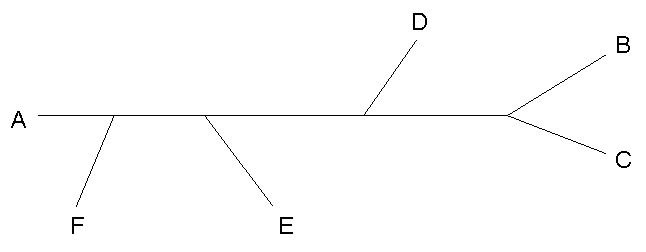 |
BCDFEA ..**** ***... ...*.* |
| Консенсусное дерево (bootstrap) | 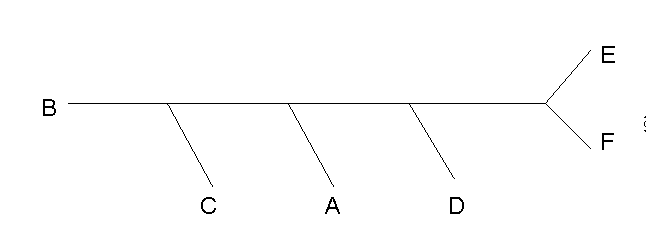 |
BCDFEA ..**** ..***. ...**. |
Полученный файл с консенсусным деревом в текстовом формате можно посмотреть здесь.
Проанализируем достоверность рекострукции при помощи бутстреп-значений ветвей консенсусного дерева. Бутстреп-значения
внутренних ветвей соответствуют числу деревьев, содержащих данную
ветвь. Чем больше бутстреп-значения, тем больше деревьев содержат
денную ветвь, значит, такая ветвь будет в консенсусном дереве.
| Консенсусное дерево | "Реальное" дерево |
Set (BCDFEA) How many times out of 100.00 |
Set (BCDFEA) How many mutations out of 100.00 |
Sets NOT included in consensus tree: |
|
Две "реальные" ветки рассматривались программой, но не вошли в
консенсусное дерево. Как видно из таблицы бутстреп-значений, ветки,
со значениями меньше где-то 40 не входят в консенсусное дерево.
Поэтому в реконструкцию вошла только одна "реально" существующая ветка.
Стоит правда отметить, что две другие ветки имеёт всё-таки не
минимальные бутстреп-значения, хотя и не близки к "порогу".
Отсюда можно сделать вывод, что данный метод можно применять только
при бОльших эволюционных расстояниях - от 40 мутаций на 100
нуклеотидов, то есть для анализа "дальних родственников", при очень же
похожих последовательностях полученное консенсусное дерево будет не
правдоподобным.
-
Изображение филогенетического дерева в графическом формате
| fdrawtree Plots an unrooted tree diagram Phylip tree file: tree_Formula.txt Phylip drawtree output file [tree_formula.fdrawtree]: tree_Formula.ps DRAWTREE from PHYLIP version 3.6b Reading tree ... Tree has been read. Loading the font ... Font loaded. Writing plot file ... Plot written to file "tree_Formula.ps" Done. |
Получившийся в результате файл имеет формат postscript и содержит графическое изображение заданного дерева:
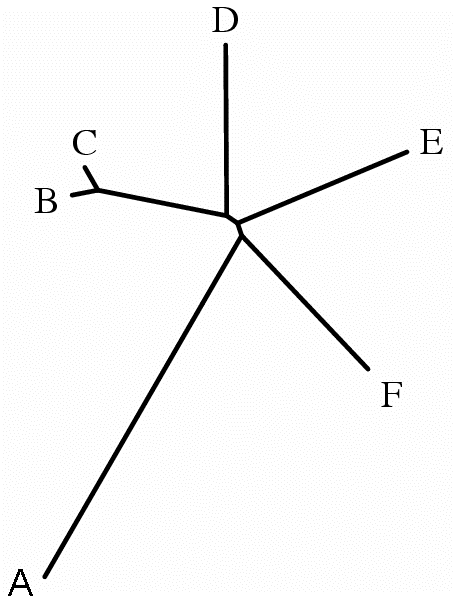
Данное дерево - неукоренённое.
Программа fdrawtree не написала букву "A" на картинке и её пришлось подрисовывать вручную.
| Исходное дерево | |
| Нарисованное программой fdrawtree | 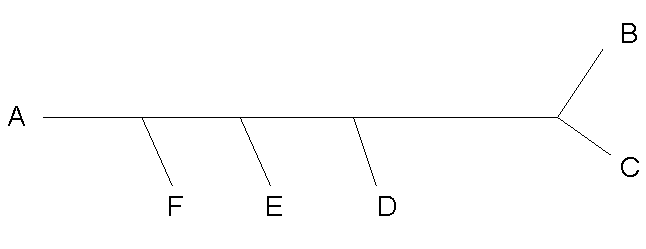 |
Было изменено положение корня при помощи изменения скобочной формулы: tree_Formula2.txt, однако на выданном программой изображении это никак не сказалось.
Дополнительное задание
- Оценить достоверность полученной топологии с помощью метода jackknife
fseqboot offsprings1.fasta -test j -auto
|
fdnaml offsprings1.fseqboot -ttratio 1 -auto
|
Далее при помощи программы fconsense получаем консенсусное дерево:
| fconsense Majority-rule and strict consensus tree Phylip tree file: offsprings1.treefile Phylip consense program output file [offsprings.fconsense]: offsprings1.fconsense Consensus tree written to file "offsprings.treefile" Output written to file "offsprings.fconsense" Done. |
Полученное консенсусное дерево:
| +--------------------A +100.0-| | | +-------------D | +-95.0-| +------| | +------E | | +-90.0-| | | +------F | | | +---------------------------C | +----------------------------------B |
Сравним полученное консенсусное дерево с исходным.
| Рисунок дерева в бескорневом виде | Описание ветвей дерева как разбиения множества листьев | |
| Исходное дерево | |
ABCDEF -**--- -***-- *----* |
| Консенсусное дерево (bootstrap) | |
ABCDEF -**--- ***--- ----** |
| Консенсусное дерево (jackknife) | 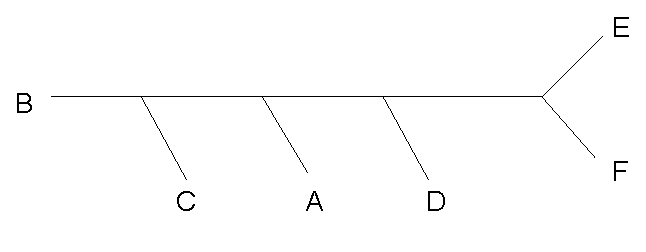 |
ABCDEF -**--- ***--- ----** |
Проанализируем достоверность рекострукции по аналогии с упражнением про бутстреп-анализ.
| Консенсусное дерево (bootstrap) | "Реальное" дерево | Консенсусное дерево (jackknife) |
Set(BCDFEA) How many times out of 100.00 |
Set(BCDFEA) How many mutations out of 100.00 |
Set(BCDFEA) How many times out of 100.00 |
Sets NOT included in consensus tree: |
|
Sets NOT included in consensus tree: |
Как видно из полученной таблицы, несмотря на сходство топологии полоченных деревьев, программы всё-таки дали различные результаты. По сравнению с бутстреп анализом, единственная совпавшая с "реальной" ветвь при использовании jackknife-анализа имеет большее бутстреп-значение, а, значит, большую статистическую достоверность. Кроме того, уменьшилось и количество рассматриваемых веток за счёт скорости алгоритма (не рассматриваются ВСЕ варианты, как в бутстреп-анализе), при этом ветки, "реально" встречающиеся всё же остались, хоть и с маленькими значениями и не внесены в консенсусное дерево, всё же рассматривались программой. Более того, jackknife приводит те же 2 "не правильные" ветви, что и бутстреп, но их "достоверность" возросла, что не очень хорошо в конкретном случае, ведь дерево неверно. Видимо, как и бутстреп, jackknife следует применять в случае бОльших эволюционных расстояний, причём при очень большом числе мутаций лучше применять jackknife из-за его большей скорости.
- Укоренение дерева в среднюю точку.
Отчёт о работе с программой fretree в текстовом формате.
Полученный в результате файл offsprings_out.treefile содержит скобочную формулу укоренённого дерева:
,-----------------------------------------------------------------------6:A |
- Изображение филогенетического дерева в графическом формате
fdrawgram -style p offsprings_out.treefile |
| -style
menu [c] Tree style output (Values:
c (cladogram (v-shaped)); p (phenogram (branches are square)); v (curvogram (branches are 1/4 out of an ellipse)); e (eurogram (branches angle outward, then up)); s (swooporam (branches curve outward then reverse)); o (circular tree)) |
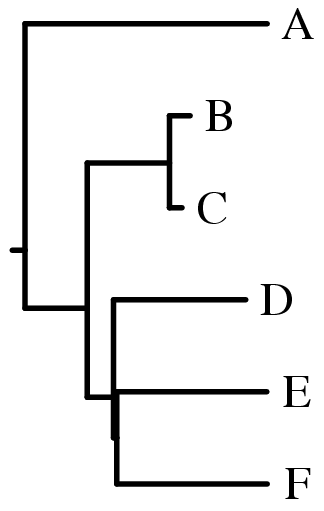 |
Полученное дерево - укоренённое, неультраметрическое, изображённое в виде филограммы, ориентированной вправо. Интересно то, что в месте пересечения веточек, несущих листья D,E и F, практически возникла трихотомия! |
Главная Первый семестр Второй семестр Третий семестр