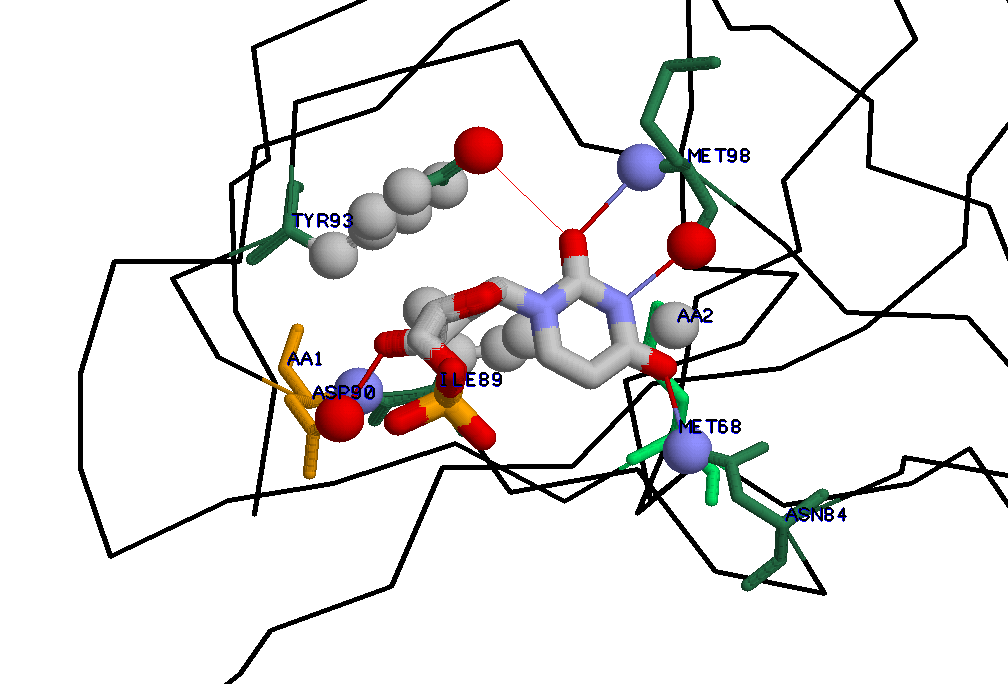
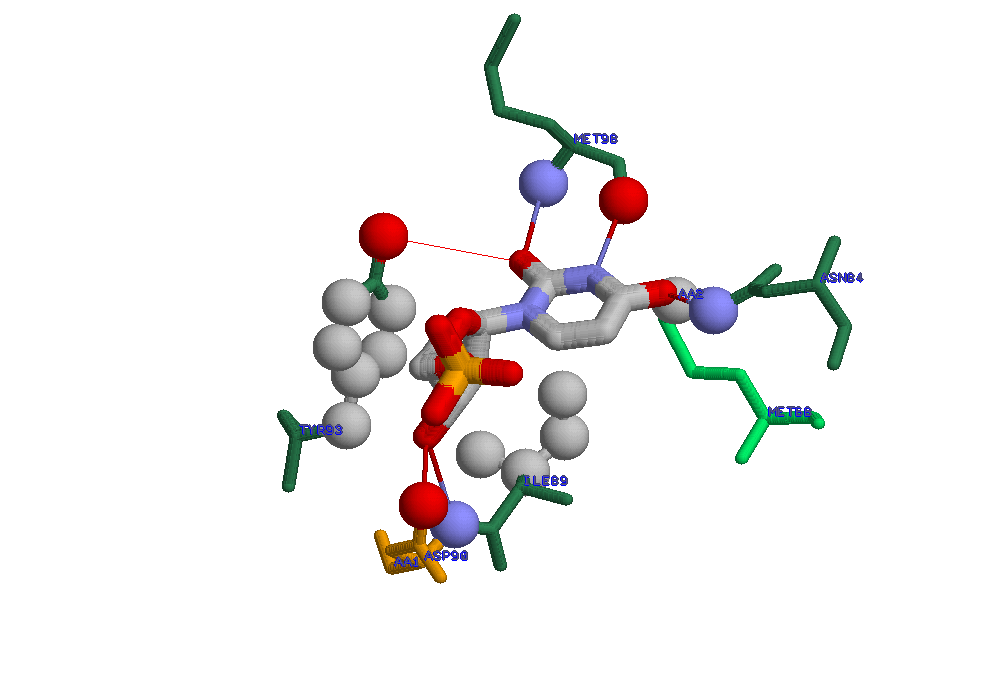
(полипептидная цепь, не участвующая во взаимодействии на рисунке отсутствует)
В генноинженерном эксперименте рассматривается возможная замена аминокислотного остатка, которая вероятнее всего
повлияет на способность белка связываться с лигандом, а также, параллельная замена, не влияющая на связывание.
Предполагается возможность возникновения таких замен при мутагенезе.
Для случая взаимодействия DUTPASE и UMP довольно сложно указать аминокислоту, участвующую в образовании водородной связи,
замена которой привела бы к потере способности белка связываться с лигандом: DUTPASE образует с UMP всего 4 водородные связи
при этом 3 из них поддерживаются за счет кислородов и азотов из остова полипептидной цепи (N и O).При замене этих аминокислот н
а практически любые аминокислоты, не очень сильно отличающиеся размером, потеря способности к связыванию не произойдет .
Предлагаемая замена: заменить аминокислоту, образующую водородную связь, за счет азота из остова белка (MET98 -AA1), на пролин:
при атоме азота, образующем в остатке пролина пептидную связь с другой аминокислотой нет водорода. Поэтому N из пролина
не способен служить донором в водородной связи.
Таким образом, в этом месте лиганд не будет связываться с белком. Если учесть что водородных связей м
ежду DUTPASE и UMP очень мало и таким образом каждая из них вносит существенный вклад в способность белка удерживать лиганд, то потеря
одной из них будет иметь большое значение.
Другой вариант, рассмотрим не столь однозначную ситуацию: заменить ASN84 (полярный) (в нем атом ND2 является донором водорода)
на гистидин (полярный ), в нем тоже есть атом азота- NE2,-который может служить донором водорода. Но боковая группа гистидина
очень громоздкая, она изменит конформацию полипептидной цепи, что может помешать связыванию.
Кроме того, донор водорода (азот)
в гистидине будет иметь другие координаты, чем соответствующий азот в аспарагине. А водородная связь очень чувствительна к направлению,
особенно к направлению донорной группы. Обычно валентная связь донора ,в данном случае N-H, прямо смотрит на тот акцептор, с коим водородная связь образуется.
Таким образом, данная замена тоже, скорее всего, повлияет на способность белка связываться с лигандом.
Замена аминокислотного остатка белка, контактирующего с UMP, но, тем не менее, предположительно, мало влияющая на способность связывания с UMP.
Заменим остаток, участвующий в гидрофобном взаимодействии с UMP -MET68(AA2) на другую аминокислоту, радикал
которой обладает гидрофобными (является неполярным) свойствами, - на изолейцин .Радикал изолейцина примерно того же размера,
что и радикал метионина .Так что, скорее всего, такая замена не повлияет на способность белка связываться с лигандом.