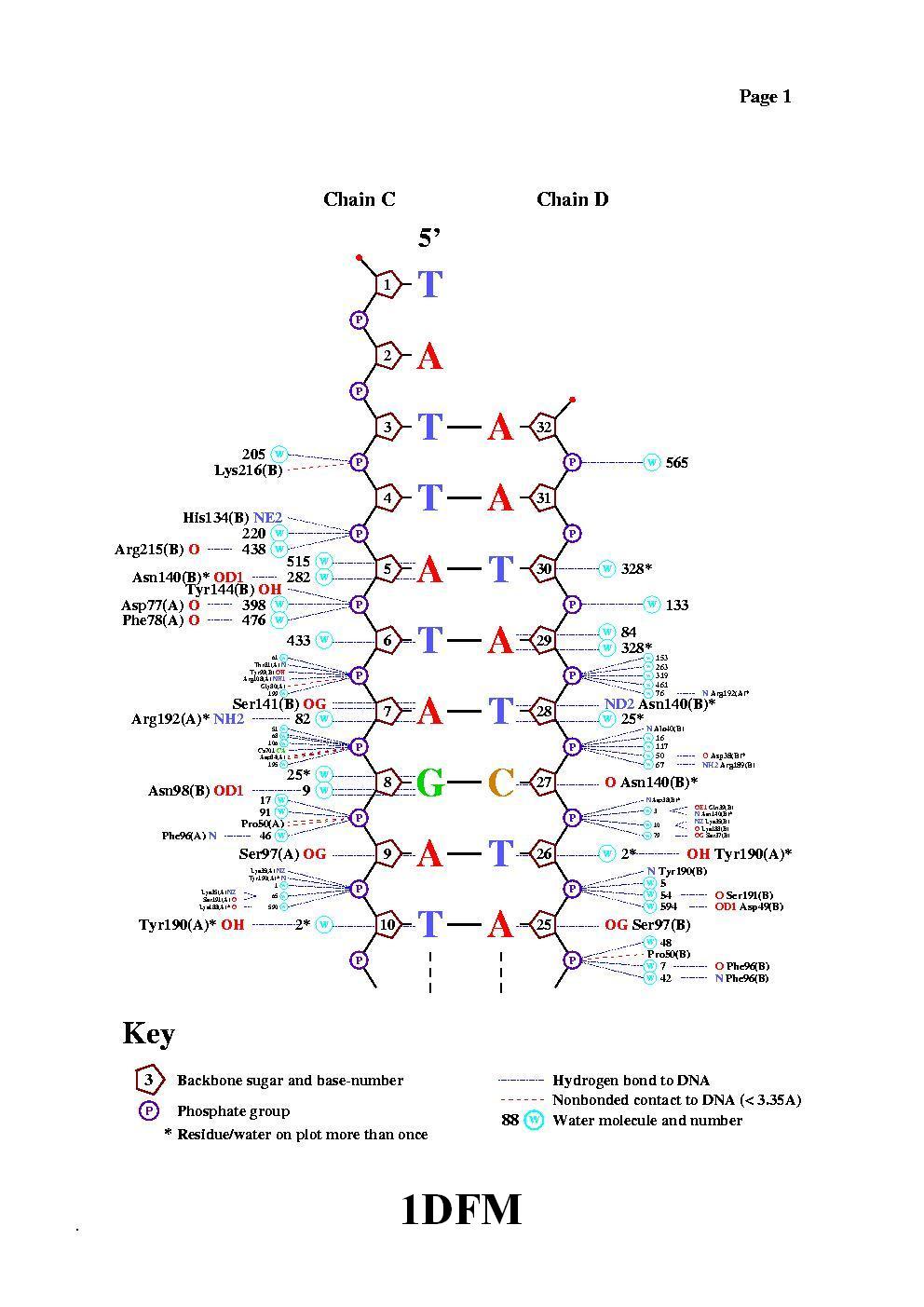
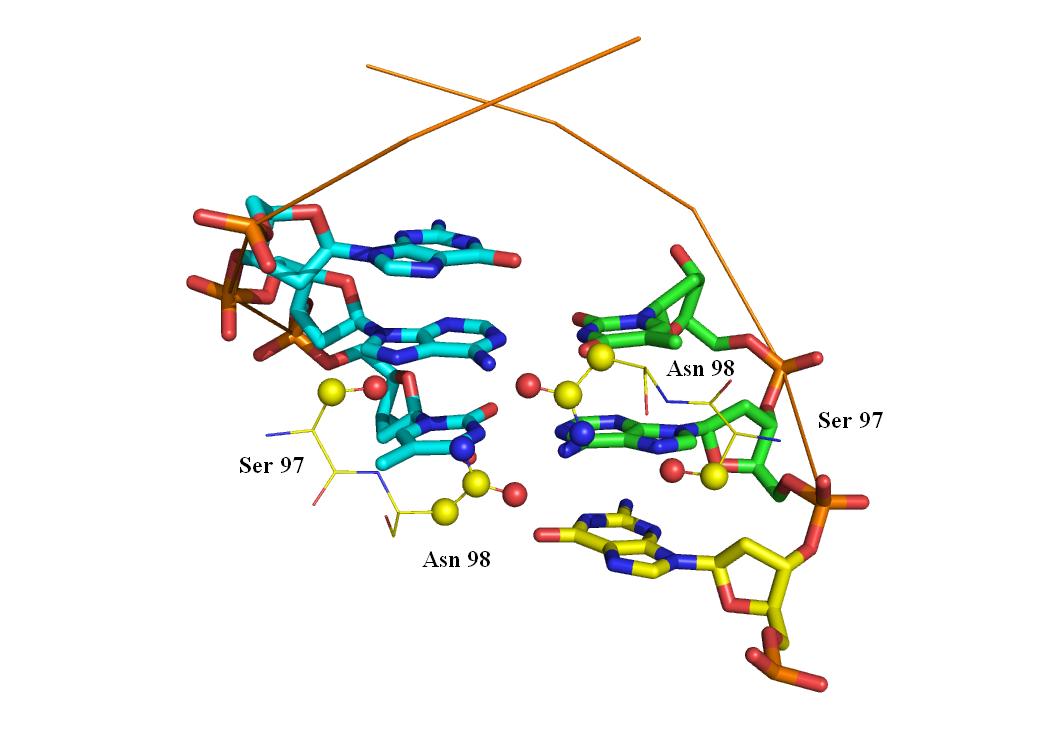
mset 1 x8
select polar_prot, ((n. n*) | (n. o*)) & !(br. (n. **') | hetatm)
select nonpol_prot, ((n. c**) | (n. s**)) & !(br. (n. **') | hetatm)
select polar_rib, (n. o3') | (n. o4') | (n. o5')
select nonpol_rib, (n. c*') & (br. (n. **'))
select polar_fosph, (n. op*)
select nonpol_fosph, (n. p)
select polar_big_way, ((n. o6) | (n. n6) | (n. n7) | (n. o4) | (n. n4)) & (br. (n. **'))
select nonpol_big_way, ((n. c5) | (n. c6) | (n. c7) | (n. c8) | ((n. c4) & ((r. *c*) | (r. *t*)))) & (br. (n. **'))
select polar_small_way, ((((n. n3) | (n. n2)) & ((r. *a*) | (r. *g*))) | ((n. o2) & ((r. *c*) | (r. *t*)))) & (br. (n. **'))
select nonpol_small_way, (n. c2) & (br. (n. **'))
distance 11, polar_rib, polar_prot, 3.5
distance 12, polar_fosph, polar_prot, 3.5
distance 13, polar_big_way, polar_prot, 3.5
distance 14, polar_small_way, polar_prot, 3.5
distance 21, nonpol_rib, nonpol_prot, 4.5
distance 22, nonpol_fosph, nonpol_prot, 4.5
distance 23, nonpol_big_way, nonpol_prot, 4.5
distance 24, nonpol_small_way, nonpol_prot, 4.5
hide
mdo 1: hide; show everything, 11 ; show ribbon, (br. (n. **'))
mdo 2: hide; show everything, 12 ; show ribbon, (br. (n. **'))
mdo 3: hide; show everything, 13 ; show ribbon, (br. (n. **'))
mdo 4: hide; show everything, 14 ; show ribbon, (br. (n. **'))
mdo 5: hide; show everything, 21 ; show ribbon, (br. (n. **'))
mdo 6: hide; show everything, 22 ; show ribbon, (br. (n. **'))
mdo 7: hide; show everything, 23 ; show ribbon, (br. (n. **'))
mdo 8: hide; show everything, 24 ; show ribbon, (br. (n. **'))
|