trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_1.pdb -skip 20
Визуализация прошла плохо. Переделаем:
trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_1.pdb -skip 20 -pbc mol
Получили файл b_pbc_1.pdb.
Результат на картинке, получила бислой:

Начнём с визуального анализа движений молекул (группа DPPC):
trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_1.pdb -skip 20
Визуализация прошла плохо. Переделаем:
trjconv -f b_md.xtc -s b_md.tpr -o b_pbc_1.pdb -skip 20 -pbc mol
Получили файл b_pbc_1.pdb.
Результат на картинке, получила бислой:
Определим площадь, занимаемую одним липидом.
Для этого получим размеры ячейки из траектории:
g_traj -f b_md.xtc -s b_md.tpr -ob box_1.xvg
В файле box_1.xvg в первой колонке содержится время, в трёх
следующих - размеры ячейки (по каждой из осей). После измерения бислоя в PyMol, определим, что
нормалью к поверхности бислоя является ось X (первая).
Построим зависимость площади, занимаемой 1 липидом, от времени в Exel:
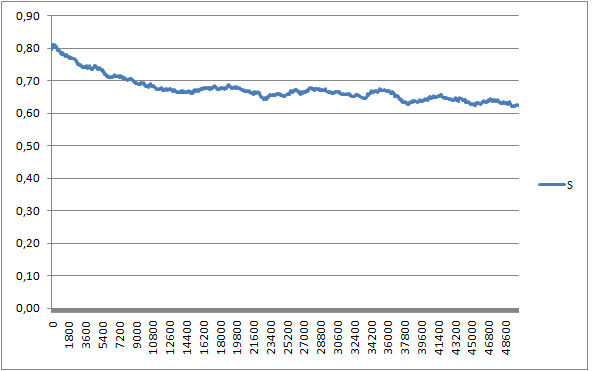
Площадь, занимаемая 1 липидом, вычисляется как произведение Y*Z, делённое на 32 (количество молекул
в монослое).
Из графика видно, что по мере формирования бислоя площадь, занимаемая 1 липидом, уменьшается до
оптимального значения 0,64 нм^2.
Определим изменение гидрофобной и гидрофильной поверхности в ходе самосборки:
g_sas -f b_md.xtc -s b_md.tpr -o sas_b.xvg
Получили файл sas_b.xvg.
В Exel построим зависимость изменения гидрофобной и гидрофильной поверхностей, доступных растворителю от времени.
Результаты на графике:
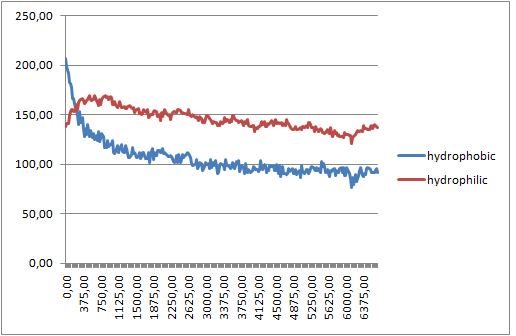
При формировании бислоя происходит уменьшение как гидрофобной, так и гидрофильной поверхности, приводящее к уменьшению энергии системы, что и служит движущей силой образования бислоя.
Традиционной мерой оценки фазового состояния бифильных молекул является мера порядка.
Для анализа нам понадобится специальный индекс файл.
Запустим анализ. Для конца траектории:
g_order -s b_md -f b_md.xtc -o ord_end.xvg -n sn1.ndx -b 45000 -d X
И для начала траектории:
g_order -s b_md -f b_md.xtc -o ord_start.xvg -n sn1.ndx -e 5000 -d X
Построим зависимости меры порядка для разных атомов липида (с головки до хвоста) на основе
выходных файлов: ord_end.xvg, ord_start.xvg.
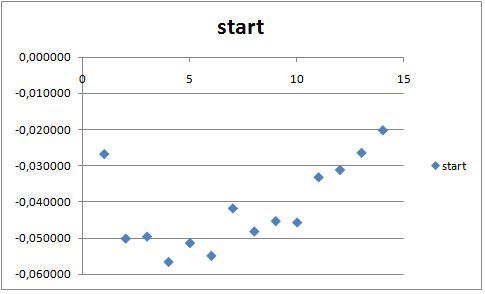
Для начала траектории:
Для конца траектории:
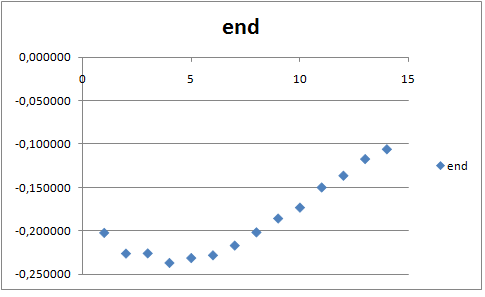
Графики имеют примерно одинаковую форму. Однако в начале траектории все значения выше, следовательно, подвижность липидов в конце траектории ниже, чем в начале, что логично, так как в бислое у липида меньше степеней свободы. На обоих графиках видно, что жирный хвост значительно подвижнее, чем гидрофильная часть, и их подвижность увеличивается к концу хвоста. Любопытно, что даже в составе бислоя хвосты могут свободно двигаться.