Получение файлов со структурами
Заданный файл PDB c ID 2IRF был получен на сайте сайта PDB, а биологическую единицу комплекса — с сайта Macromolecular Structure Database EBI. В результате поиска по PDB-идентификатору выдача содержала только одну модель
В результате обе структуры оказались совершенно одинаковыми.
И вылядят следующим образом:
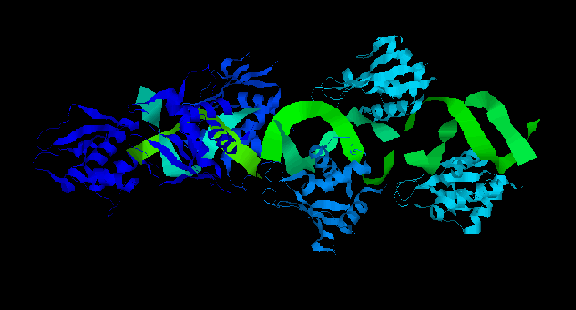
Как видно, структура не симметрична, и поэтому биологическая единица оказалась идентична структуре, содержащейся в PDB-файле.
Исследование контактов между молекулами белка и нуклеиновой кислоты
Таблица. Контакты разного типа в комплексе 1IRF
| Полярные | Команда, с помощью которой получены данные | Гидрофобные | Команда, с помощью которой получены данные | Всего | |
| Контакты белка с ... | |||||
| ... остатками 2'-дезоксирибозы | 14 | select within(3.5, *.O?* and dna) and polar |
68 | select within(4.5, *.C?* and dna) and hydrophobic |
82 |
| ... остатками фосфорной кислоты | 81 | select within(3.5, *.O?P and dna) and polar |
48 | select within(4.5, *.P and dna) and hydrophobic |
129 |
| ... остатками азотистых оснований со стороны большой бороздки | 15 | select within(3.5, (oxygen, nitrogen) and mjg and base) and polar |
10 | select within(4.5, carbon and mjg and base) and hydrophobic |
25 |
| ... остатками азотистых оснований со стороны малой бороздки | 0 | select within(3.5, (oxygen, nitrogen) and mig and base) and polar |
0 | select within(4.5, carbon and mig and base) and hydrophobic |
0 |
Как видно из таблицы, самое большое количество контактов как полярных, так и гидрофобных белка с остатками фосфорной кислоты. Это вполне закономерно, так как молекула ДНК не устойчива, и эти контакты несут функции поддерживания остова молекулы. Следующие по числу контактов контакоты белка с остатками остатками 2'-дезоксирибозы. Эти контакты так же несут функции поддерживания структуры ДНК. Примечательно, что контактов белка остатками азотистых оснований со стороны малой бороздки совсем нет. Возможно, это обусловлено тем, что малая бороздка достаточно глубока и при этом узка (в исследуемом pdb-файле ДНК находится в А-форме) для полярных или гидрофобных контактов. Большая бороздка в свою очередь достаточно широка, чтобы могли быть возможны полярные или неполярные контакты. И эти контакты белка с остатками азотистых оснований со стороны большой бороздки несут функции узнавания ДНК белком. Именно среди этих контактов следует искать остатки связывания ДНК.
Для получения данных использовался скрипт P:\y05\Term3\dna.def, определяющий три нужных множества:
base — все атомы оснований ДНК (эквивалентно "dna and not backbone");
mjg — атомы оснований ДНК, обращённые в большую бороздку;
mig — атомы оснований ДНК, обращённые в малую бороздку.
Полярными атомами ДНК считались все атомы кислорода и азота, а неполярными — все атомы углерода.
Поиск специфических контактов, обеспечивающих узнавание сайта в молекуле ДНК
Специфические контакты узнавания ДНК белком сожержатся среди контактов белка с остатками азотистых оснований со стороны большой бороздки, так как таковых нет с остатками со стороны малой бороздки.Атомы белка, участвующих в контактах с остатками азотистых оснований со стороны большой бороздки:
| Полярные контакты | Гидрофобные контакты |
CB CYS 2983 chain L SG CYS 2983 chain L NH1 ARG 2082 chain L NZ LYS 2075 chain L NH1 ARG 882 chain K CB CYS 683 chain J SG CYS 683 chain J NH1 ARG 682 chain J SG CYS 483 chain I NH1 ARG 482 chain I CB CYS 283 chain H SG CYS 283 chain H NH1 ARG 282 chain H NZ LYS 275 chain H CE LYS 275 chain H |
CA ALA 2084 L N ALA 2084 L CB ALA 2079 L CB ALA 684 J CA ALA 684 J N ALA 684 J CB ALA 679 J CA TRP 258 H CA ALA 284 H CD ALA 279 H |
Подробнее был рассмотрен контакт между атомами азота ARG882K.N и G1023B.N7.
Изображение данного контакта:
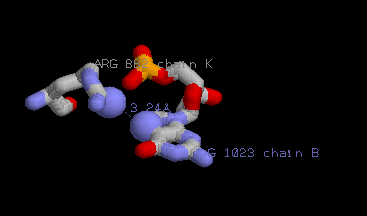
Азотистое основание и аминокислотный остаток подписаны. Более крупно выделены взаимодействующие атомы.
Описание функций исследованного белка
Описание белка IRF2_MOUSE в UniProt
Функции: Специфически связывается с регуляторноными областями генов
интерферона I и интерферон-зависимыми MHC класса I (the interferon
consensus sequence (ICS)), расположенными в 3'-5' направлении,
и подавляет эти гены.
Субъединица: Взаимодействует с BRD7 (по сходству).
Внутриклеточное положение: Ядро.
Стимулирование: Вирусами и IFN.
Сходства: Пренадлежит семейству IRF.
Сходства: Несет 1 ДНК-связывающий домен из 5ти повторяющихся триптофанов.
Описание соответствующего ДНК-связывающего домена в Pfam
Домен IRF находится в БД PfamA. Pdb-файл 2irf.pdb содержит структуру комплекса irf-2 и ДНК.
| Домен | Цепь белка | Начальный остаток | Конечный остаток |
| IRF | G | 5 | 113 |
| IRF | H | 205 | 313 |
| IRF | I | 405 | 513 |
| IRF | J | 605 | 713 |
| IRF | K | 805 | 913 |
| IRF | L | 2005 | 2113 |
Описание домена IRF из БД Pfam.
Транскрипционный фактор интерферон-регуляторного фактора.
Это семейство транскрипционных факторов важно в регуляции ответа интерферонов на
инфицирование вирусом и в регуляции интерферон-зависимых генов.
3 из 5 консервативных триптофановых остатков связываются с ДНК.
Описание из БД INTERPRO (запись IPR001346)
Экспрессия генов интерферона I (α и β интерфероны) запускается многими агентами,
включая атаку вирусами. Связывание интерферон-регуляторного фактора 1 (IRF-1) с областями,
называемыми interferon consensus sequence (ICS), расположенными в 3'-5' направлении на интерфернционных генах,
является связующим звеном в стимулирование выработки интерферона I.
Другие факторы также могут связываться с областью ICS, включая IRF-2, который не функционирует как
активатор, но скорее подавляет функции IRF-1 под влиянием определенных обстоятельств.
IRF-белки несут консервативные области N-конца около 120 аминокислот в длину,
которые сворачиваются в структуру, которая связывается специфически с областями ICS;
остальные же части последовательности варьируют зависимо от определенных функций белка.
Изображение выравнивания домена IRF:
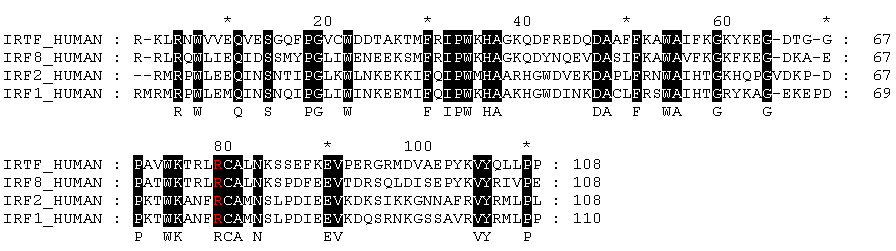
Консервативные на 100% участки покрашены черным. Остаток, исследовавшийся в п. 3, выделен красным на черном фоне. Этот остаток оказался консервативным на 100%, то есть можно надеяться, что выбранный остаток участвует в специфическом взаимодействии с молекулой ДНК.
Раскрашенное выравнивание, созданное с использованием Jalview (Michele Clamp)
{kind=link}
Построение диаграммы контактов белка с ДНК
Использовалась программа nuclplot командойnucplot 2IRF.pdb
Полученная диаграмма
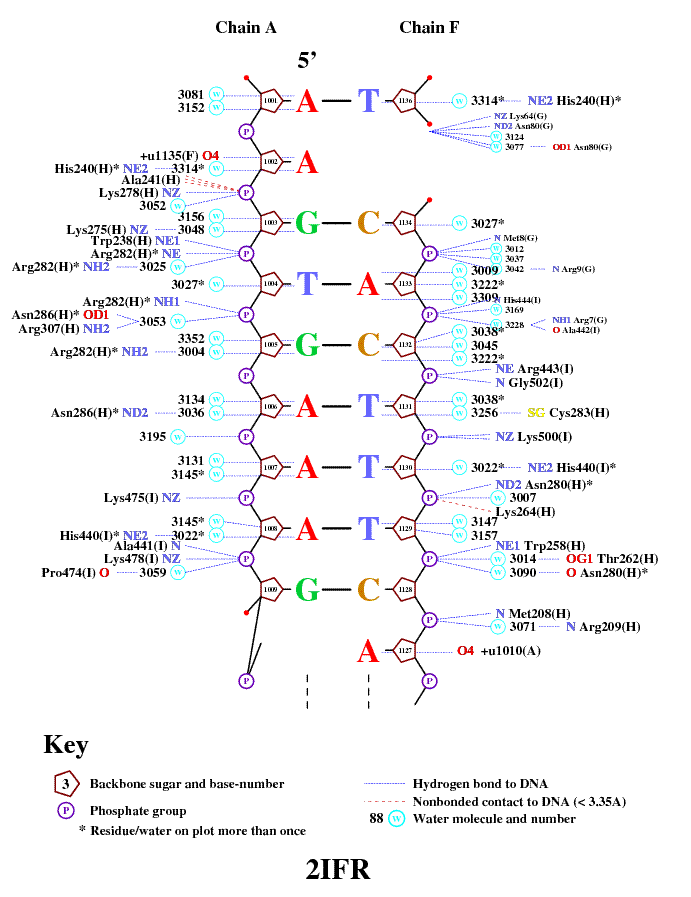
В файле 2ifr.bond, созданным программой, были найдены следующие данные о взаимодействии атомов остатка ARG 882 K:
Donor Acceptor Distance ARG K 882 NH1 G B 1023 O2P 2.77 ... ARG K 882 NH2 HOH 3005 O 2.72 ... HOH 3005 O G B 1023 O6 2.68Как видно, NH1 ARG 882 chain K является донором во взаимодействии с атомом O2P G 1023 chain B. То есть предположение, что данный атом участвует в специфическом связывании ДНК, было ошибочным, но тем не менее атом азота NH1 ARG 882 chain K участвует в неспецифическом "поддерживающем" связывании с остатками фосфорной кислоты ДНК.
В случае с остатком ARG 882 chain K специфическое взаимодействие происходит через молекулу воды HOH 3005. Атом NH2 ARG 882 chain K взаимодействует с атомом O HOH 3005, являясь донором, а атом O HOH 3005 в свою очередь является донором во взаимодействии с атомом O6 G 1023 chain B. Достаточно интересное взаимодействие. То есть впринципе можно считать, что остаток ARG 882 chain K участвует в специфическом взаимодействии с молекулой ДНК.
Данные были взяты из файла 2ifr.bond, так как на диаграмме изображена "эквивалентная", но не "та самая" цепь белка.