
При детальном рассмотрении нашлась одна пара и одна тройка атомов белка, взаимодействующих с общим атомом ДНК. Как взаимодействующий среди них был выбран атом, образующий самую короткую связь, а остальные на рисунке обведены и в дальнейшем в учет не брались. В итоге остались 9 пар взаимодействующих атомов:
| Контакты белка с ... | Полярные | Гидрофобные | Всего |
| остатками 2'-дезоксирибозы | - | - | 0 |
| остатками фосфорной кислоты | 7(голубой) | - | 7 |
| остатками азотистых оснований со стороны большой бороздки | 1(сине-зеленый) | 1(фиолетовый) | 2 |
| остатками азотистых оснований со стороны малой бороздки | - | - | 0 |
Очевидно, взаимодействий очень мало, особенно если учесть, что эти данные лишь предположения реальных контактов. Однако, чтобы найти наиболее вероятные остатки, отвечающие за спецефичность, необходимо ещё сильнее уменьшить полученную выборку.
При рассмотрении структуры белкового участка вблизи контакта с ДНК, было найдено, что большинство найденных ранее атомов принадлежит остаткам, располагающимся в α-спирали(остальные в неструктурированных участках). Эта спираль образована десятью остатками с 76-го по 85-й и располагается в большой борозде ДНК, что, видимо, обеспечивает очень прочное связывание молекул:

Вид на α-спираль сверху. Бирюзовые шарики - взаимодействующие атомы ДНК. Специфичные остатки белка показаны в проволочной модели
Удалив из внимания остатки не входящие в эту α-спираль, мы получили 6 остатков с претензией на спецефичность: глутаминовая кислота 77, лизин 78, серин 80, аргинин 80, аргинин 81, глицин 82. Из них первые четыре взаимодействуют с остатками фосфорной кислоты, а сами их контакты располагатюся закономерно и симметрично (выделены на рисунке ниже), что добавляет уверенности в их спецефичности:

α-спираль сбоку. Остальной белковый остов не показан. "Красные" остатки спирали не участвуют в узнавании.
Что же касается двух остатков, взаимодействующих с атомами большой бороздки (81 и 82), то их об их специфичности трудно сказать что-то определенное, однако, возможно, они (или один из них) выполняют роль цепи, за которую α-спираль втягивается внутрь большой бороздки, где и закрепляется при помощи других остатков.
Для отдельного рассмотрения был выбран остаток серин 80, взаимодействующий с остатком фосфорной кислоты цитозина 7:
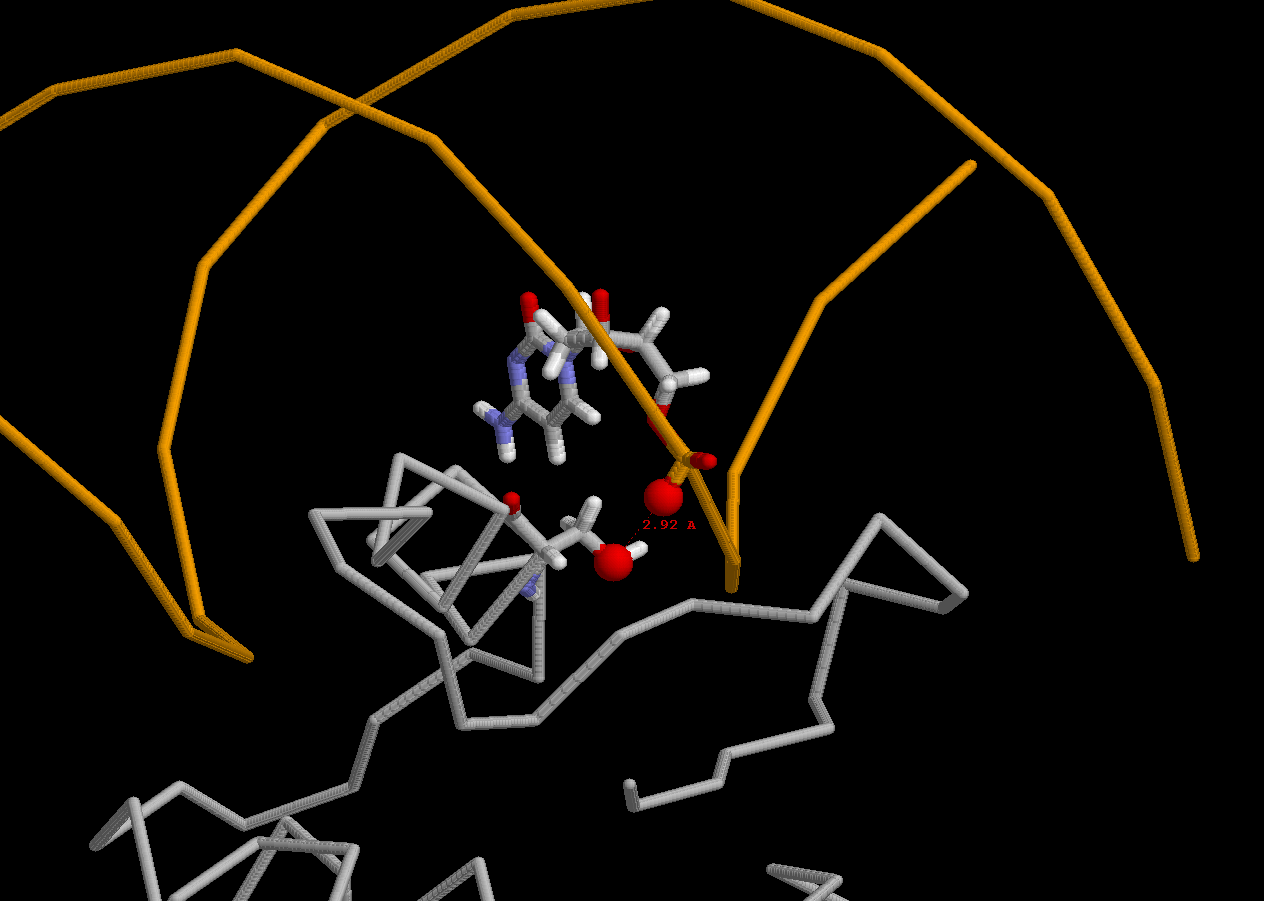
Рассматриваемый аминокислотный участок принадлежит белку ETS1 из Homo Sapiens, являющимся транскрипционным фактором. Отвечает за деформацию эритробластов и фибробластов. Может принимать участие в патогенезе моноклеточной лейкемии человека.
Факторы транскрипции — молекулы белка, которые связываются с определенными последовательностями ДНК в геноме, приводя к индукции или запрещению генной транскрипции. Ets oncogene является подобным участком. Его длина равна 85-90 аминокислот, и он известен как ETS (эритробласт определенное преобразование) область. Эта область богата положительно-заряженными и ароматическими остатками, и связывается с богатыми пурином участками ДНК. Область ETS была идентифицирована в других участках транскрипции, типа PU.1, human erg, human elf-1, human elk-1, GA белок, и множество других. Обычно она находится в C-конце белка, за исключением ELF-1, ELK-1, ELK-3, ELK-4 и ERF, где она найдена в N-конце. Анализ ЯМР структуры областей Ets показал, что они содержат три альфа-спирали и четыре раза переплетённые бета-тяжи. Третья альфа-спираль способна связываться с большой бороздкой ДНК. Различные члены семейства белков Ets показывают высокую специфичность к ДНК. Области Ets и аминокислотные последовательности белков влияют на специфичность. Изменение единственной аминокислоты в области Ets может изменить специфичность ДНК.
Для проверки специфичности было проведено выравнивание выборки из 15-ти доменов:

Участок α-спирали выделен красным. Видно, что серин 80 был выбран не очень удачно. Более (а исходя из данной выборки - абсолютно) консервативны
оказались аргинины 81 и 84, а вот глицин 82 совершенно неконсервативен. Таким образом можно оставить 4 остатка: 78, 80, 81 и 84.
В конце концов нам осталось доверить всю работу программе nucplot. Она выдала очень "сухую схему":

Как ни странно, но по иронии, единственным "надежным" контактом nucplot выбрала именно остаток серина 80, который мы "случайно" выбрали выше! Все четыре
возможно специфичных контактов подчеркнуты салатовым. Фиолетовым подчеркнут 77-й остаток, который плохо выравнивается, но тем не менее отмечен nucplotом.
©Nenchuk Spartak, 2007