на главную
назад
Исследование белок-нуклеиновых контактов
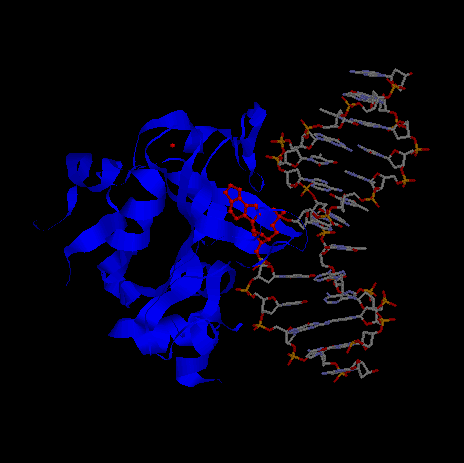 |
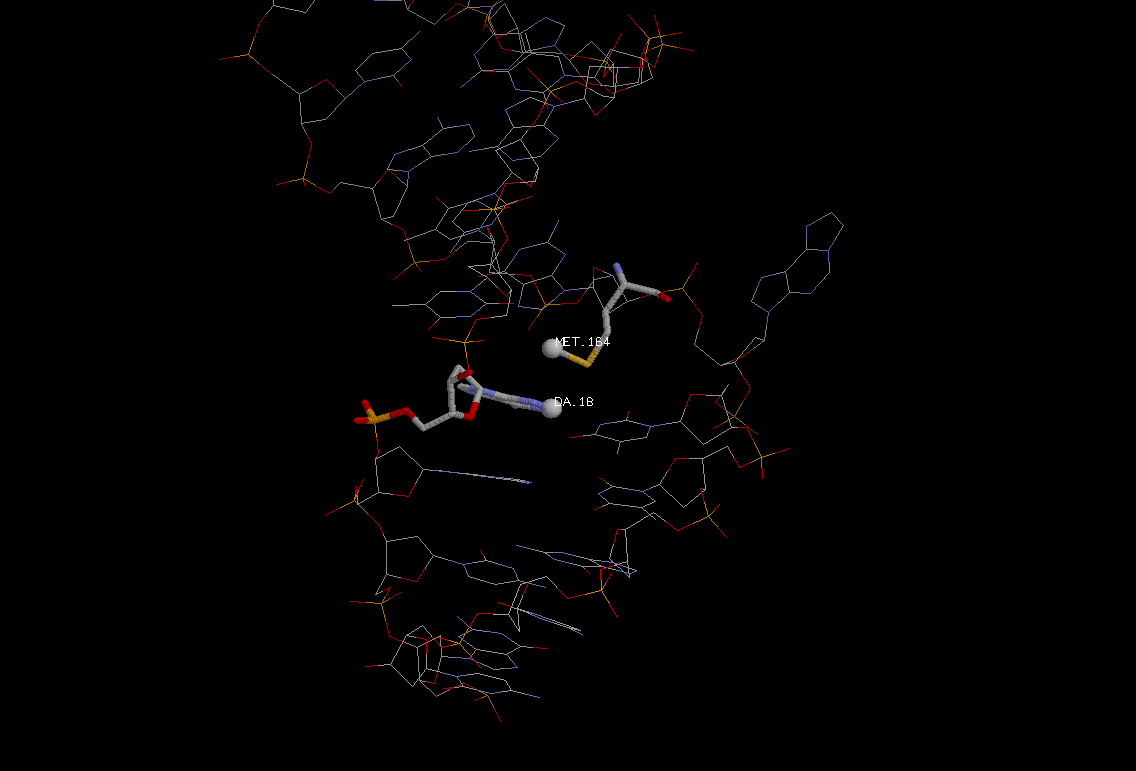
- Синим покрашен белок.
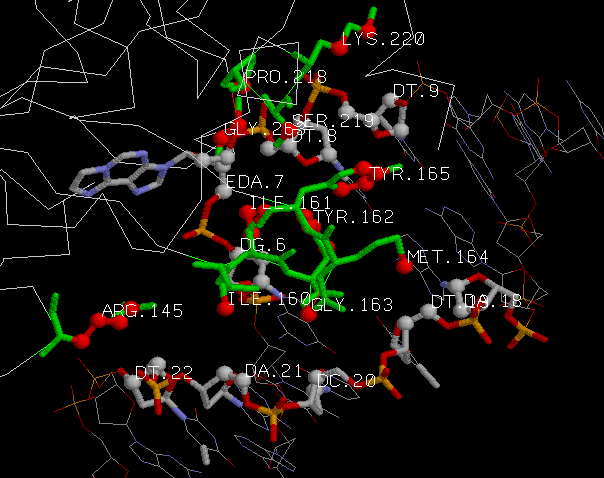
- Красным - атомы, определенные Rasmol'ом как лиганды. Причем туда входит остаток модифицированного аденина (ЭТАН-АДЕНИН) из цепи ДНК, развернутый ОТ спирали.
К лигандам также относится ион Na(обозначен красным шариком, расположенным отдельно).
|
Исследование контактов между молекулами белка и нуклеиновой кислоты
Исследование проводилось с помощью команды Rasmol.
Был использован скрипт dna.def, который выделял нужные множества (mjg - множество атомов оснований, обращенных в большую
бороздку, mig - в маленькую бороздку). Был написан скрипт script.txt, с помощью которого и был осуществлен поиск
контактов.(скрипт работает для не изменённого файла 1EWN.pdb)
Таблица. Контакты разного типа в комплексе ДНК - 3-метил-аденин ДНК гликозилаза
| |
Полярные |
Гидрофобные |
Всего |
| Контакты белка с ... |
|
|
|
| ... остатками 2'-дезоксирибозы |
4 |
24 |
28 |
| ... остатками фосфорной кислоты |
12 |
--- |
12 |
| ... остатками азотистых оснований со стороны большой бороздки |
0 |
5 |
5 |
| ... остатками азотистых оснований со стороны малой бороздки |
1 |
1 |
2 |
| ... c модифицированным основанием (этан-аденином) |
4 |
24 |
28 |
Для подробного описания полученных результатов сначала разберемся в функционировании данного фермента.
3-метил-аденин ДНК гликозилаза участвует в репарации(base-excision repair), "очищает" молекулу ДНК от измененных оснований, которые мешают нормальной
экспрессии гена. Она находит модифицированные азотистые основания, а именно - алкилированные (напр. этан
аденин), "выворачивает" их из спирали ДНК (основание связывается с активным центром, по-видимому, с натрием), а затем
проявляя N-гликозидную активность отщепляет основание от соответствующего сахара. При этом освободившееся место занимается другим основанием,
посредством другого фермента.
Легко понять, что исследуемая структура представляет практически конечную стадию работы фермента (нуклеотид уже развернут). Этим можно легко объяснить то, что имеется
большое количество взаимодействий белка с ЭТАН-АДЕНИНОМ, который уже прочно связался с ферментом и готов отщепиться от сахара.
Фермент должен прочно крепиться к ДНК во время реакций. Большое количество контактов белка с остатками дезоксирибозы и фосфорной кислоты
как раз и делает фермент прочно связанным с молекулой ДНК. Эти контакты сконцентрированны вокруг развёрнутого модифицированного основания.
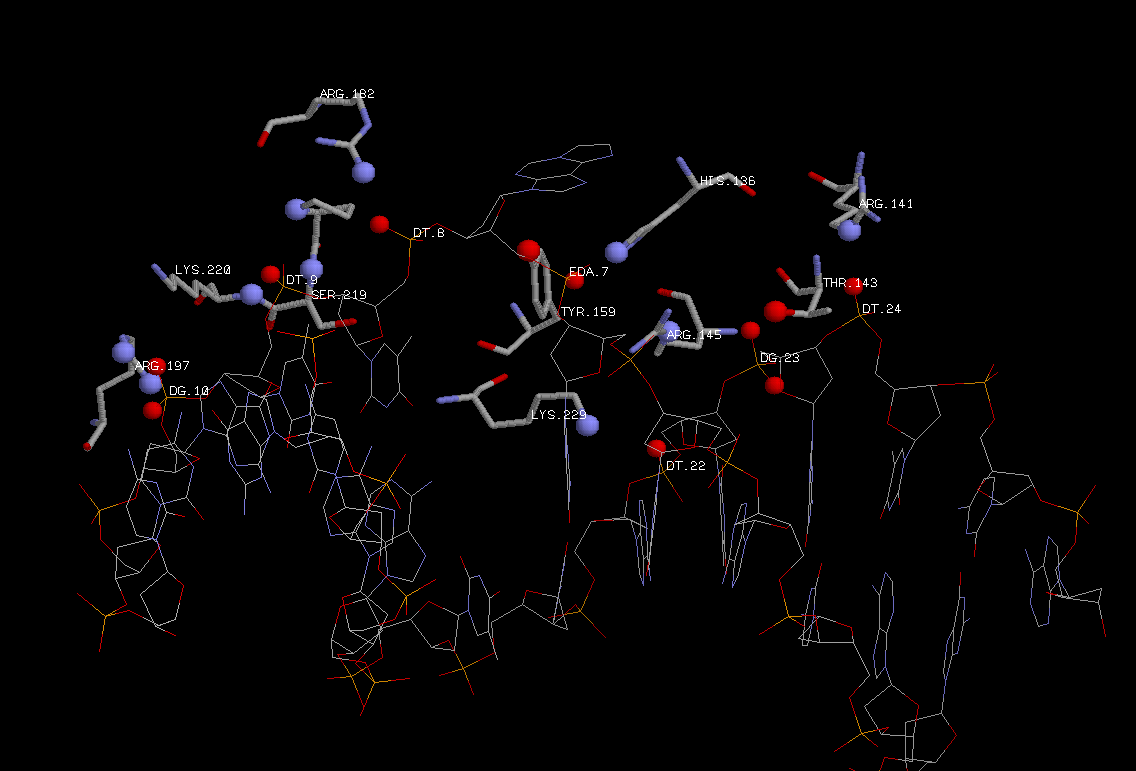
А вот что касается контактов с большой и малой бороздкой, тут все по-иному. Большое количество контактов такого рода, возможно, имело место
на ранних стадиях работы фермента, когда он еще "искал" этан-аденин или начинал с ним связываться. Теперь же контактов таких немного из-за ненадобности. Белок изменил
свою конформацию и изменил её в ДНК, тем мамым потерялись старые связи. Оставшиеся взаимодействия способствуют связыванию фермента с ДНК и развороту этан-аденина (
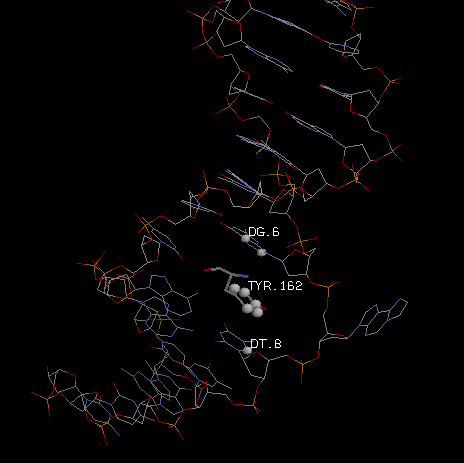
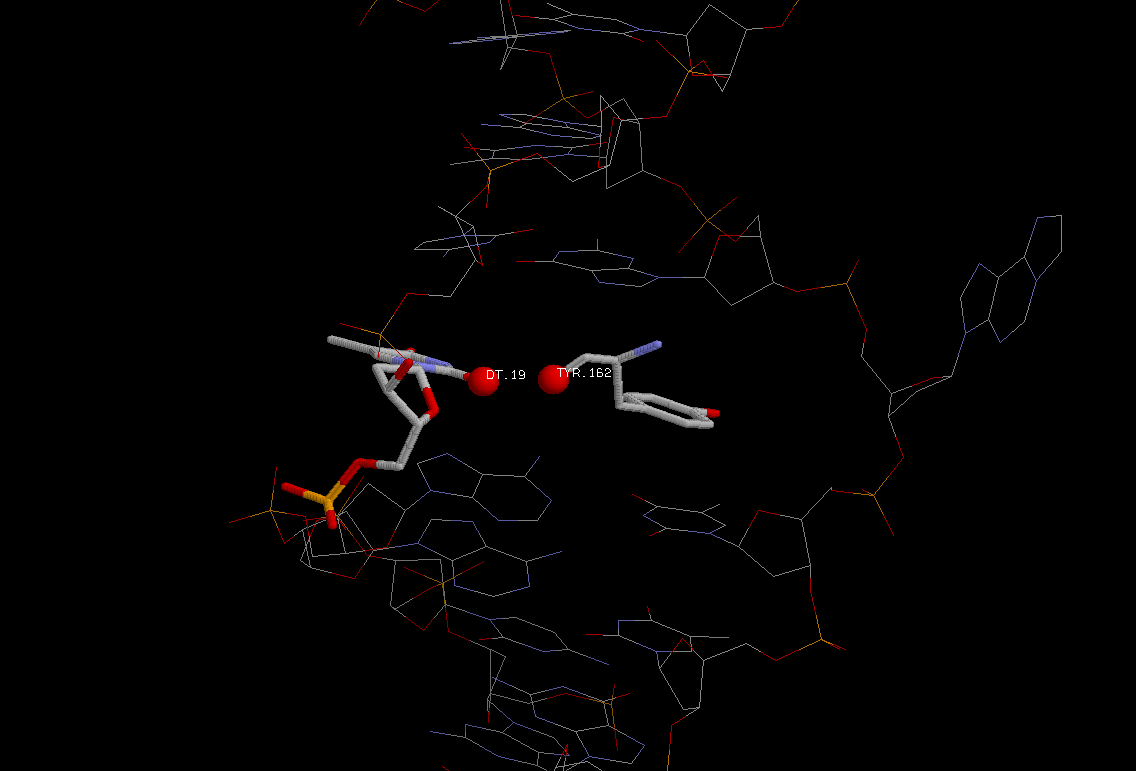
на место колец основания встало кольцо тирозина,
связанного гидрофобно с атомами большой бороздки).
Специфические контакты.
На самом деле предположить, какие аминокислотные остатки данного фермента отвечают за специфичность
узнавания ДНК, довольно сложно, т.к. представленная структура, как уже говорилось, показывает одну из последних
стадий ферментативной реакции. Но даже по найденным взаимодействиям можно кое-что сказать.
Так как данный фермент должен "узнать" модифицированный ацилированный остаток ДНК, он обязан иметь
остатки аминокислот, реагирующих с "лишними" атомами углерода (присоединённые в результате ацилирования) гидрофобными взаимодействиями.
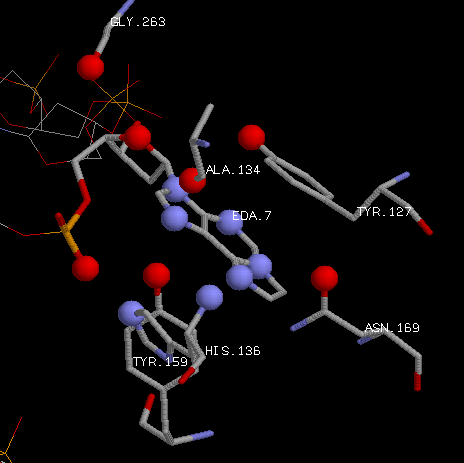
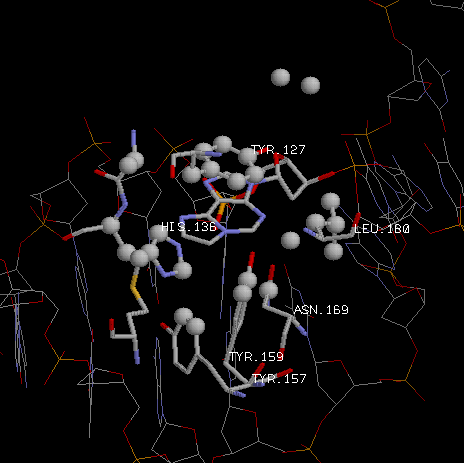
Посмотрим на гидрофобные взаимодействия этан-аденина и аминокислот. |
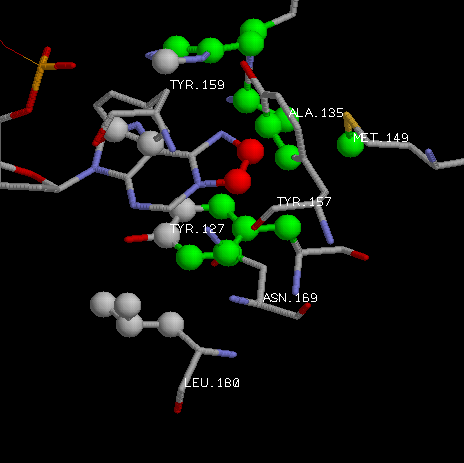 |
| Красным выделены атомы углерода присоединившегося этана. А зеленым - атомы углерода аминокислотных
остатков, способных к гидрофобному взаимодействию с ними.
Поэтому можно сказать, что аминокислотные остатки TYR-157, ALA-135,
TYR-127, ASN-169, MET-149 (в большей степени конечно аланин и тирозин-127 из-за большего кол-ва контактов)
отвечают за специфичное узнавание белком модифицированного основания. Тирозин-127 вообще образует гидрофобное
стекинг-взаимодействие с кольцом этан-аденина.
|
Описание белка UniProt
- Название белка - DNA-3-methyladenine glycosylase
- Имя в банке UniProt - 3MG_HUMAN
- Идентификатор - P29372
- ДНК-связывающий домен : цепь А , с 87 по 284 аминокислоту (Pur_DNA_glyco)
-
Cellular component nucleoplasm
Molecular function damaged DNA binding
Molecular function identical protein binding
Biological process base-excision repair
Biological process DNA dealkylation
Подробнее о функции белка было сказано выше.
Приведем выравнивание Pfam ДНК-связывающих доменов гомологичных белков разных организмов.

Взглянув на выравнивание, можно заметить, что многие позиции в домене полностью консервативны. Данные позиции скорее всего определяют
специфичность узнавания ДНК.
Красными точками отмечены позиции, которые по предположению задания 3 отвечают за специфичное узнавание белком ДНК.
Положения 127, 157, 169 - полностью консервативны. Другие же 2 позиции тоже довольно консервативны, но в меньшей степени.
Эти данные могут послужить подтверждением того, что предсказанные аминокислоты, отвечающие за специфичность узнавания, действительно являются таковыми.
Файл полученный с помощью программы nucplot
(использован изменённый файл 1EWN3.pdb)
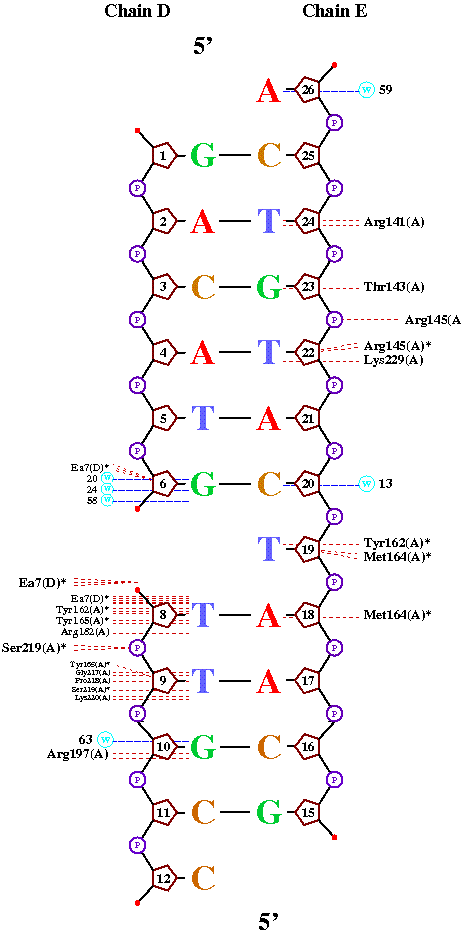 |
Интересно отметить, что данная программа не указывает гидрофобных контактов с белком, но зато показывает не учтенные нами в во
втором задании взаимодействия с молекулами воды, которые играют довольно значительную роль в формировании структуры.
В целом то что показал nucplot соотносится с тем, что мы получили используя RASMOL. Много совпадающих контактов. НО все же в RASMOL'e взаимодействий, даже полярных было найдено больше.
|
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}