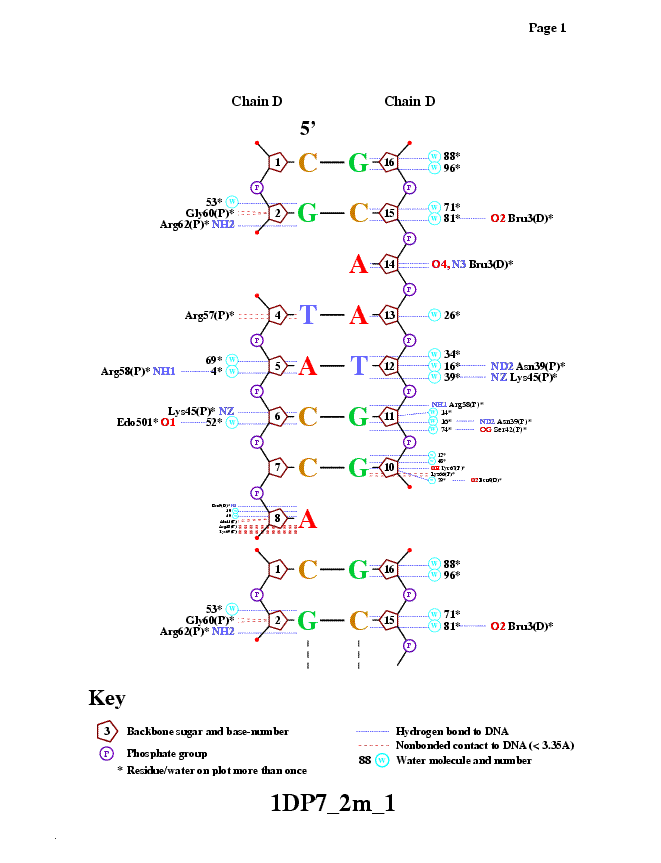
| Полярные | Гидрофобные | Всего | |
| Контакты белка с ... | |||
| ... остатками 2'-дезоксирибозы | 6 | 44 | 50 |
| ... остатками фосфорной кислоты | 24 | - | 24 |
| ... остатками азотистых оснований со стороны большой бороздки | 10 | 10 | 20 |
| ... остатками азотистых оснований со стороны малой бороздки | 2 | нет | 2 |
|
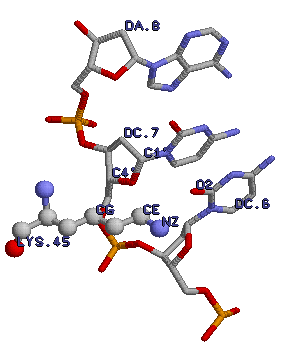
Здесь представлены остаток лизина (lys45) и 6-7 нуклеотиды цепи ДНК. Подписаны атомы, вовлеченные во взаимодействия, описанные выше (расстояния между соответствующими атомами не больше 3,5 Å для полярных контактов, не больше 4,5 Å для гидрофобных). |