Файл с нужной структурой, скачанный с сайта PDB, называется 1EMH.pdb.
В записи полученной структуры обозначения нуклеотидов не однобуквенные, а двубуквенные: DA,DT, DG, DC (D=дезокси-). Rasmol эту запись не ассоциирует с множеством ДНК, поэтому приходится с помощью команды define доопределять нуклеотиды в множество dna1. К нему же я отнесла P2U - 2'-дезокси-псевдоуридин-5'-монофосфат. Нумерация атомов в рибозе, остатке фосфорной кислоты и основаниях стандартная.
Из-за вышеописанного обозначения нуклеотидов в структуре для использования скрипта dna.def названия оснований в нем также были переобозначены. Псевдодезоксиуридин не относится ни к одной бороздке, т.к. смотрит вбок, а не внутрь спирали. Отмечу, что не все атомы углерода, которые по рассказанному на лекции материалу были отнесены к большой или малой бороздке, в данном скрипте определены как принадлежащие одной из бороздок. Соответственно, они не включаются в определенные ниже полярные и неполярные группы.
С помощью команды define были определены полярные и неполярные множества (см. скрипт).
В пределах 3,5 А от кислородов дезоксирибозы оказалось 2 атома кислорода (атомы углерода я сразу исключила из запроса). Рибоза образует 2 полярные связи. В качестве возможных атомов белка, образующих гидрофобные связи с ДНК, я рассматривала только атомы углерода (причина очевидна), что было отражено в запросах. В пределах 4,5 А был обнаружен 21 атом углерода, возможно образующий гидрофобные связи с дезоксирибозой.
Было найдено 3 атома кислорода и 4 атома азота. Неполярных контактов фосфорная кислота не образует (нет углеродов).
Не было найдено ни одной полярной связи этой группы с белком, но есть 2 полярных связи атомов азота оснований с кислородами воды, находящейся внутри молекулы белка. Неполярных связей 3.
Не было найдено ни одной полярной связи этой группы с белком, но нашлось 3 связи азотов оснований с кислородом воды. Была найдена одна неполярная связь.
Найдено 7 атомов, с которыми возможны полярные связи: 5 азотов и 2 кислорода. Также найдена одна полярная связь с кислородом воды и 24 неполярных связи псевдоуридина и белка.
Написан скрипт, который позволяет посмотреть каждое вышеобозначенное множество.
Таблица. Контакты разного типа в комплексе 1EMH
| Полярные | Гидрофобные | Всего | |
| Контакты белка с ... | |||
| ... остатками 2'-дезоксирибозы | 2 | 21 | 23 |
| ... остатками фосфорной кислоты | 7 | 0 | 7 |
| ... остатками азотистых оснований со стороны большой бороздки | 0 | 3 | 3 |
| ... остатками азотистых оснований со стороны малой бороздки | 0 | 1 | 1 |
| ... остатком псевдоуридина | 6 | 24 | 30 |
Из таблицы видно, что основания со стороны бороздок образуют небольшое число гидрофобных связей. Связей, образованных атомами малой бороздки меньше, возможно, из-за того, что она менее доступна для молекул белка. В любом случае, эти связи не играют значительной роли. Гораздо больше связей образует дезоксирибоза, почти все они гидрофобные; несколько полярных связей образуют остатки фосфорной кислоты. Но наибольший вклад в образование комплекса вносит, конечно же, обращенный наружу псевдоуридин.
Очевидно, что в качестве специфичного в данном случае стоит рассматривать контакт белка с дезоксиуридином.
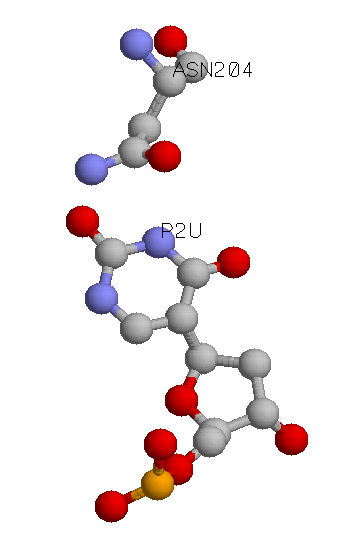
Основание контактирует с несколькоми аминокислотными остатками белка: ASN204A, PHE158A, ASP145A. Наиболее интересным мне показался остаток ASN204A, т.к. с ним нуклеотид может образовать 2 полярные связи.
Урацил-ДНК-гликозилаза удаляет из ДНК остатки урацила, разрезая N-гликозидную связь. Урацил в ДНК может появляться в результате неправильного присоединения dUMP остатков ДНК-полимеразой или деаминизирования цитозина. Последовательность урацил-ДНК-гликолазы очень консервативна. За активность этого белка отвечает аспарагиновая кислота в положении 154 (в структуре pdb она имеет номер 145), ее замена приводит к потери активности. Аспарагин в 213 положении отвечает за специфичность, его замена на аспарагиновую кислоту приводит к цитозин-ДНК-гликозилазной активности. Это как раз указанная мной выше аминокислота asn204a. Замена тирозина 156 на аланин дает тимин-ДНК-гликозилазную активность.* На сайте Pfam было сделано выравнивание, по которому видно, что аспарагин ASN204A, выбранный мной, консервативен.
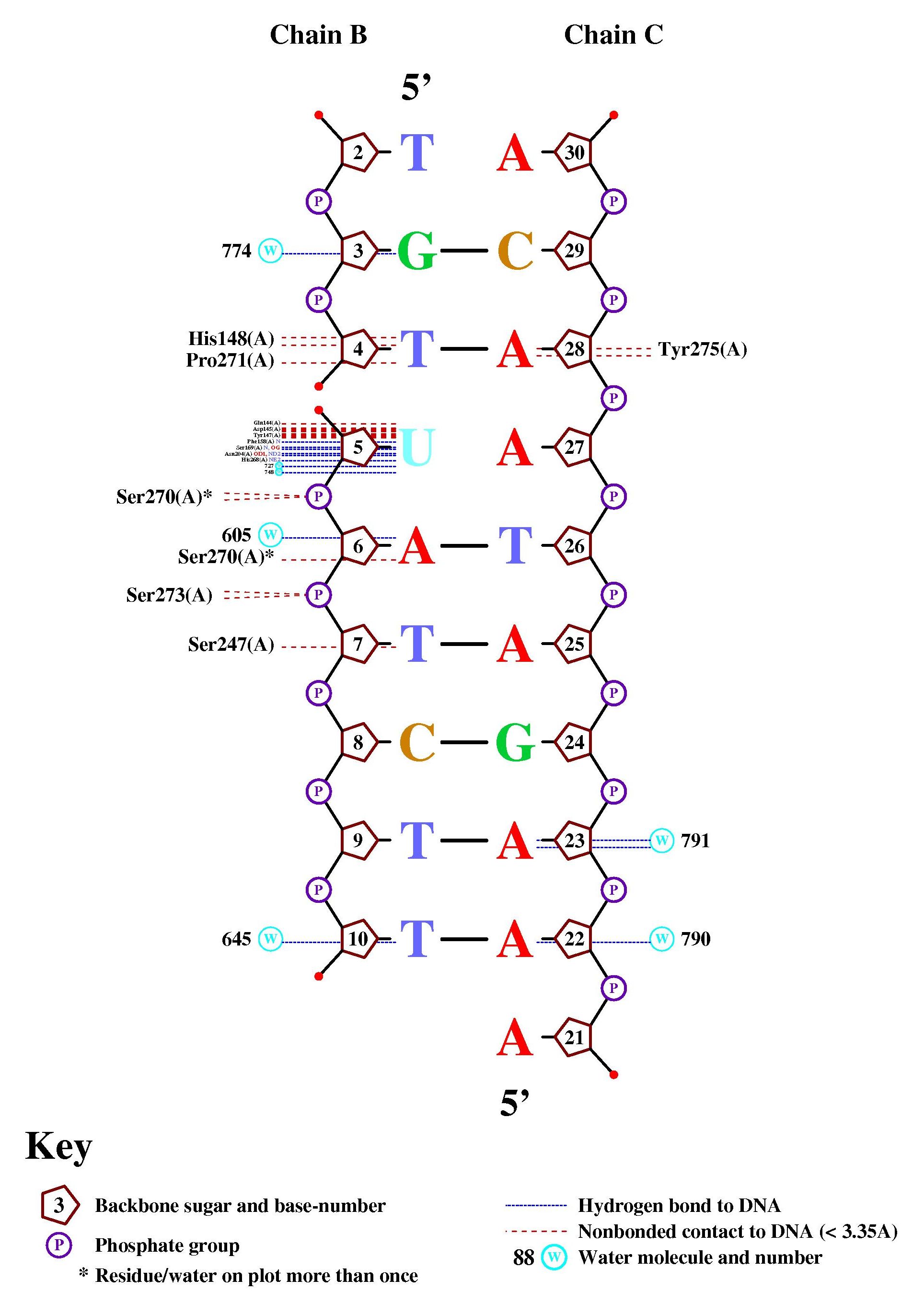
С помощью программы nucplot, были найдены следующие контакты (псевдоуридин обозначен как уридин 5):
Найдено 4 водородных связи с остатком фосфорной кислоты, как и в моей таблице. Найдено несколько полярных связей оснований, что отличается от результата анализа трехмерной структуры. С остатком псевдоуридина я нашла больше связей при изучении пространственной структуры, чем через nucplot. Разница в результатах возможна из-за различных алгоритмов нахождения связей. Но из результатов анализа программы также можно сделать вывод, что псевдоуридин играет ключевую роль в узнавании фрагмента ДНК белком.
* Информация взята с сайта Pfam и базы данных UniProt.